 HUN-REN Szegedi Biológiai Kutatóközpont
HUN-REN Szegedi Biológiai Kutatóközpont 

Minden eukarióta sejt legdinamikusabb alkotórésze az aktin monomerekből felépülő, lineáris polimerek által alkotott mikrofilamentum hálózat. Az elmúlt évtized kutatásai jelentősen megváltoztatták az aktinnak a sejt életében betöltött szerepéről addig kialakult képet, és teljesen új szempontokkal gazdagították azt. Az egyik ilyen új felfedezés, az aktin sejtmagi jelenléte, csak a közelmúltban vált egyértelművé. Mára világossá vált, hogy a sejtmagban az aktin olyan alapvető folyamatok résztvevője, mint a génátírás és annak szabályozása, az RNS képzése és exportja, a kromatin szervezése, a DNS hibajavítás vagy éppen a sejtmag membrán felépülése.
Kutatócsoportunk egyik fő érdeklődési területe a sejtmagi aktin biológiai jelentőségének vizsgálata, és a sejtmagi jelenlétét biztosító mechanizmusok felderítése. Az utóbbi évben elkezdtük a sejtmagi aktinnak az immunsejtek differenciálódásban betöltött szerepét is tanulmányozni. Ezekhez a kísérletekhez a kiváló modellszervezetet, a Drosophila melanogastert használjuk.
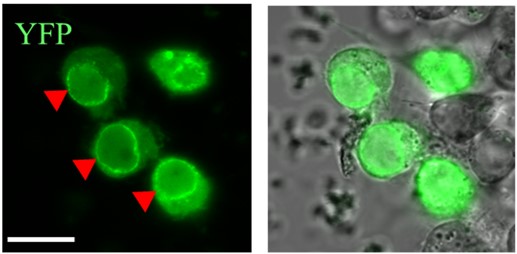
Az aktin és a Ketel nevű sejtmagi importin közötti fizikai kapcsolatot igazolja a zöld fluoreszcens jel a sejtekben.
Csoportunk az aktinkötő ERM fehérjék sejtmagi funkcióinak kutatásával is foglalkozik. Az aktin citoszkeleton nagyszámú és igen fontos feladatainak végrehajtásában legalább 80, különböző aktinkötő fehérje működik közre a citoplazmában. Ezek közé tartoznak az aktinkötő Ezrin-Radixin-Moesin (ERM) fehérjecsalád tagjai, amelyek membránfehérjéknek az aktin hálózathoz való horgonyzásával az aktin dinamika fontos szabályozói a sejtben. A három paralóg ERM gén gerincesekre jellemző, míg más fajokban, például a Drosophilában csak egyetlen ERM gén található. Az ERM fehérjék alapvető szerepet játszanak a sejtadhézióban, sejtmozgásokban, a sejten belüli szállítási folyamatokban, s így végső soron kulcsszereplői a sejtpolaritás, morfogenezis és tumor metasztázis folyamatainak.
A Drosophila egyetlen ERM képviselőjének, a Moesin vizsgálatával bizonyítottuk, hogy az ERM fehérjék is jelen vannak a sejtmagban. Meghatároztuk a Moesin sejtmagba jutását szabályozó fehérjemotívumokat és mechanizmusokat, vizsgáltuk a sejtmagi importjának dinamikáját, és szabályozását. Felderítettük, hogy a Mediátor komplexszel együttműködve részt vesz a hősokk gének működésének szabályozásában, illetve az mRNS exportban.
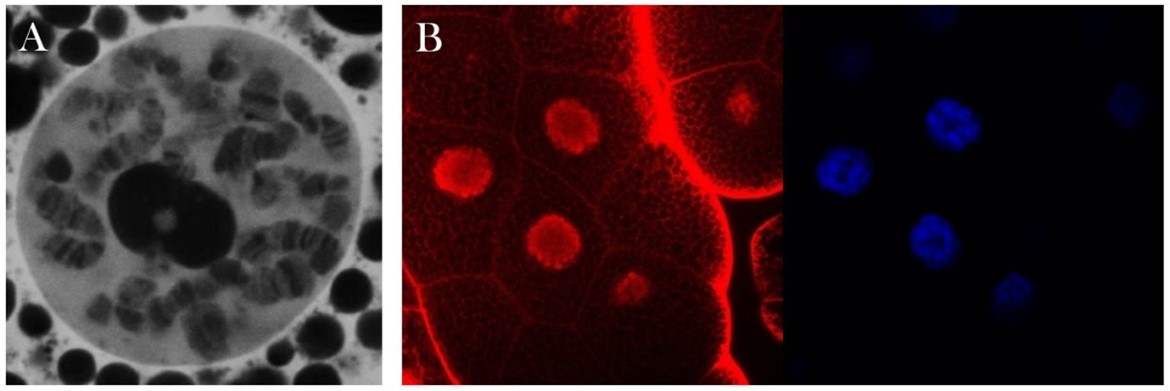
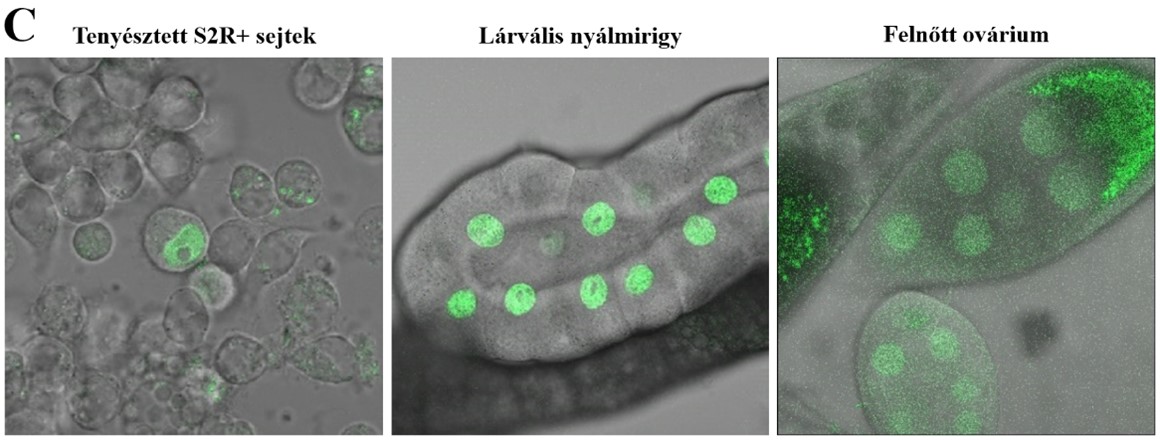
A Drosophila Moesin fehérje sejtmagi lokalizációja. A) A Moesin sejtmagi eloszlása (fehér) egy interfázisos politén sejtmagban. B) A Moesin (piros) halmozódása a sejtmagokban (kék – DAPI). C) A moesin és a Mediátor komplex közötti fizikai kapcsolatot a sejtmagi zöld fluoreszcencia igazolja.
Egy Moesin mentes sejtmagokkal rendelkező mutáns létrehozásával meghatároztuk a fehérje sejtmagi lokalizációjának biológiai jelentőségét és a sejtmagi hiányából adódó problémákat. Ezek a kísérletek vezettek arra a felismerésre, hogy a sejtmagi Moesin a hősokk indukálta chaperon gének kifejeződéséhez is szükséges.
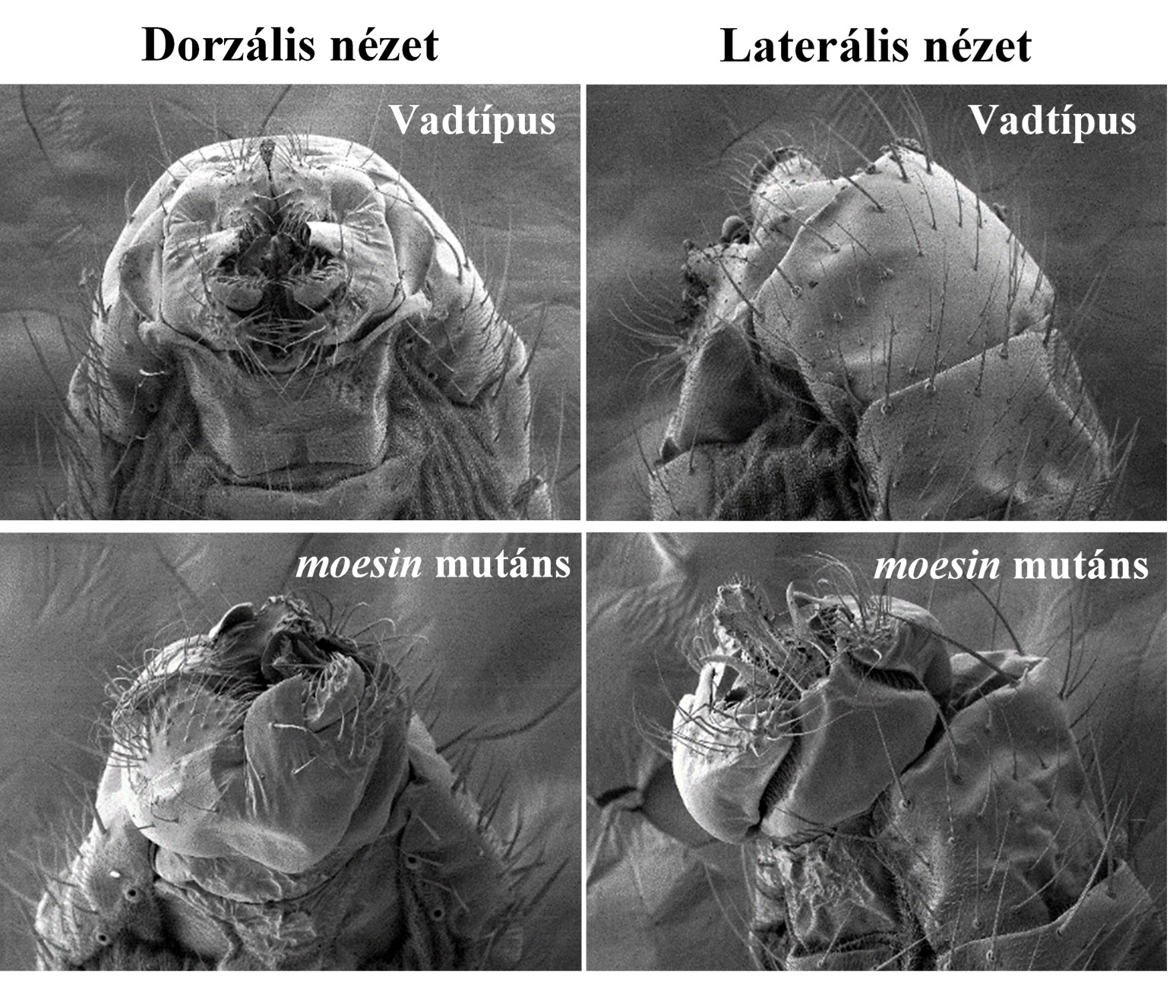
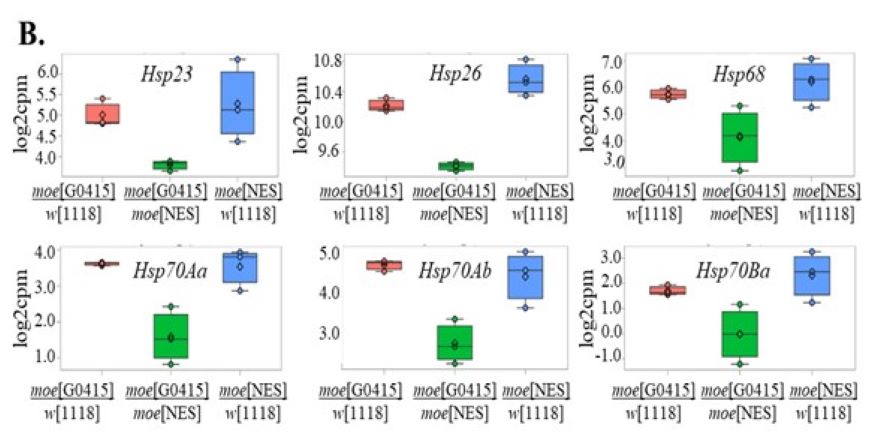
A) A moesin mentes sejtmagokkal rendelkező mutáns hímek ivarszerve elfordul fejlődés közben. B) A mutánsokban a hősokk chaperonok expressziós szintje alacsony (zöld).
A vizsgálatokhoz korszerű fénymikroszkópos technikákat, biokémiai és molekuláris biológiai módszereket, valamint az ecetmuslicát és tenyésztett Drosophila sejtvonalakat egyaránt használunk.

tudományos főmunkatárs

tudományos munkatárs

tudományos munkatárs

tudományos munkatárs
ügyvivő szakértő
|
VILMOS Péter
|
tudományos főmunkatárs | publikációk | CV |
|
KRISTÓ Ildikó
|
tudományos munkatárs | publikációk | CV |
|
SZABÓ Anikó
|
tudományos munkatárs | publikációk | CV |
|
KOVÁCS Zoltán
|
tudományos munkatárs | publikációk | CV |
|
ABONYI Csilla
|
ügyvivő szakértő | publikációk |